|
私達,ホモ・サピエンスは約20万年前にアフリカの北東部に誕生しました.その名は「知恵のある人」という意味です.人類は世界中に居住するなど,世界最強の動物として発展してきました.しかし,これは決して単純な発達ではなく,繁栄・衰退を伴うものでした.これに大きく影響を与えたのが気候・環境変動だったと考えられていますが,現在のところその証拠は極めて不十分です.
2. 人類が変えだした地球環境「私達の未来を占う環境学」
人類活動は20世紀,ついに自然の地球表層システムに影響を与えるほど大きなものとなり,地球環境問題がクローズアップされています.二酸化炭素は温室効果気体なので地球温暖化をもたらすとともに,酸性化気体なので,大気中の濃度の上昇は海洋のpHを下げ,有孔虫,サンゴ,貝などの炭酸カルシウム殻を溶解させます.このため,これらの生物は今後急速に衰退していくと予想されています.私達は,精密飼育実験などを通じて,人間が変え始めた地球環境が及ぼす影響を評価するとともに,過去の酸性化を復元し,人類の将来について考えます.(Inoue, Hayashi, Ishikawa, Mishima, Ozaki, Mori, Suzuki, Nagoshi)
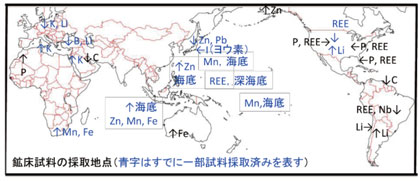
3. 大規模元素濃集体の限界に関する研究「資源と地球システム」
地球システムには,しばしば限界が存在します.例えば,観測された最大の地震はM9.5のチリ地震で1,000km長の断層が動きました.M11の地震の場合,2万 kmが一度にずれ動かねばならないので「たぶん起こらない」とされます.地球上に存在する元素濃集体(資源)の規模にも限界が存在します.それは地質学的,地球化学的,鉱物学的な制約によって規定されているからです.本研究では「地球史上最大規模の鉱床型」に焦点を絞ることで,元素濃集プロセスの主要支配要因,そして,規模を最終的に制限する因子を解析し,地球システムの構成要素がもつ限界を明らかにします.(Asakura)
本研究は2015年FYに本格的に開始予定です.
 |
|
|
| 現在,「資源」と「環境」は別物として扱われることが多いのですが,両者は「ひと繋がり」です.陸上風化,火山活動に伴う熱水活動からの元素の供給,最終的に濃集体形成に至る一連のプロセスとなっているからです.Fe, Mn, C(石炭,石油)などの堆積性鉱床は「地球環境の大変化により作られました」.このプロセスを,次世代型同位体(Li, B, Ca, Mg, Si,Fe, Zn, I, Sr(87Sr/86Srでなく88Sr/86Sr))と特定有機化合物の分析・解析による明らかにします.下が課題例です.(Wang) |
|
 |
4. 次世代同位体が拓く地球システムにおける物質循環の研究
|
|
|
| 元素には同位体が存在します.元素の化学的性質は陽子数できまり,同じ元素で中性子の数が異なるのが同位体です.放射壊変しないものは安定核種,するものは放射性核種と呼ばれます.自然界では核種ごとに反応速度,化学結合のしやすさ,拡散や蒸発などにわずかな差が存在します.そこで,化学反応の前後で同位体存在比が変化し,これは同位体分別と呼ばれます:(1)相対質量差(Dm)が大きいほど,大きな分別が起こります.(2)同位体交換反応は通常,温度の上昇とともに減少します(概して1/T2に比例).(3)重い同位体は強い結合(結合の弾性定数が大きい)に濃集します.(4)平衡状態にある物質間の結合強度に差があるほど分別が大きくなります.軽元素(H,C,O,Sなど)は同位体分別が大きく測定が容易なため,半世紀以上にわたり大きく発展してきました(出版された炭酸塩のδ18Oの論文は1万本以上). |
|
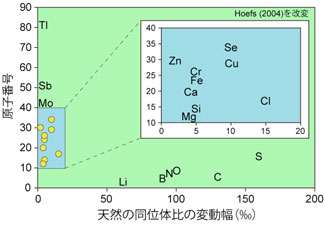 |
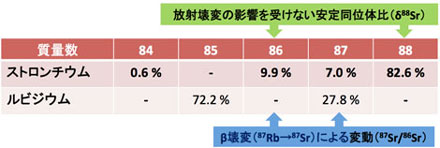
21世紀にはいりマルチコレクタ型ICP-MS(誘導結合プラズマ質量分析計)が発達し,測定精度が一桁以上向上したので,次世代型同位体の研究が急速に進展しています(地化学試料のMg同位体の論文は現時点で約100本,88Sr/86Srではわずか11本,鉱体のLi同位体に関する論文は私達のAraoka et al. (2014)が初めてです(3.図 参照)).これから発展するであろう次世代型同位体(Li, B, Ca, Mg, Si,Fe, Zn, I, Sr(87Sr/86Srでなく88Sr/86Sr))をさまざまな物質循環のプロセスの解明に利用していきたいと考えています.
<δ7Li>
Li同位体分別は大きいので,水・岩石反応の指標として近年注目されています.島弧系熱水のLi濃度,同位体は0.34-1.30 mmol/kg,+4.3 to +7.2 ‰でした.堆積物に被覆された熱水では3.40-5.98 mmol/kgと増加し,δ7Li比は+1.6 to +2.4 ‰と低くなりました.(Arakoka,Hirashima)
<δ44Ca>
Caは石灰化プロセスをトレースするのに非常に役立ちます.造礁サンゴの精密飼育実験によるとδ44Caの温度依存性は0.02 ‰/°Cで,無機的に沈殿したものと温度依存性は類似しているものの系統的に約 +0.4 ‰のずれが見いだされました.これはCa2+が海水からサンゴに取り込まれる際に生物学的に生じた同位体分別であると考えられています.(Inoue)
5. 陸域の水環境「アジアの水環境」
|
|
|
<ヒマラヤ山系の河川>
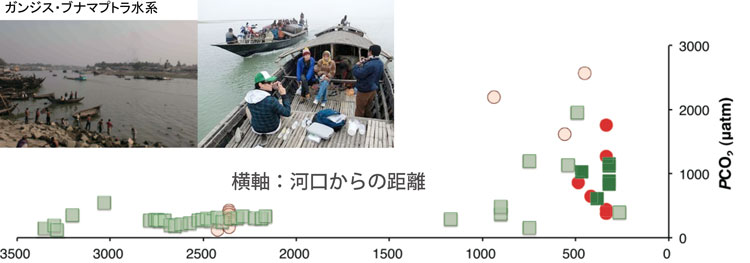
南・東南・東アジアは,活発な化学風化+物理風化で特徴づけられます.新生代後期のヒマラヤ造山運動はpCO2の減少に重要な役割を演じたとの有名な仮説があります.バングラデシュ,ミャンマー,タイ,ベトナムでサンプリングを行いました.従来ケイ酸塩風化の寄与が大きいと考えられてきましたが,炭酸塩風化が卓越していることが明らかとなり上記仮説と逆の結果が得られました.
|
|
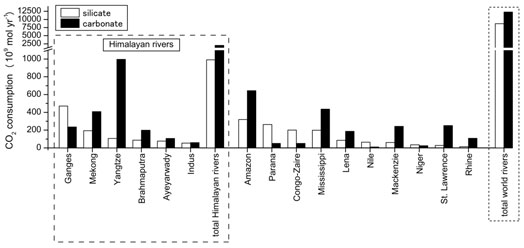 |
PCO2は,上流高所(河口からの距離が長い)では化学風化により小さくなりますが,低地では土壌中の有機物分解により数千ppmに達し,河川から大気に二酸化炭素が放出されていることを示していました.将来の温暖化への呼応では, 化学風化よりも生物活動がより敏感に反応すると考えられ, それに伴いPCO2が増加し,河川はCO2の大気への供給源として機能し得ると予想されます.(Manaka, Otani, Ushie, Suzuki)
<河川からの元素の流入>
河川は海洋への元素の主たる供給源です.これまで元素の滞留時間が(=海洋での現存量/河川流入量)濃度のみで計算されてきましたが,河川の同位体を分析し,連立方程式を解くことで,より正確に評価できます.Li, B, Ca, Mg, Sr, Siなどの次世代型同位体を分析・解析中です.(Osawa)
6. 海底熱水鉱床と海洋「新ホモール計画(IODP)」
|
|
|
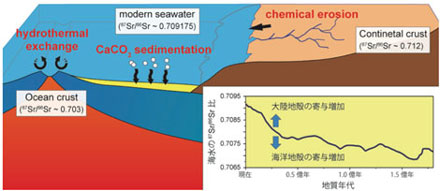
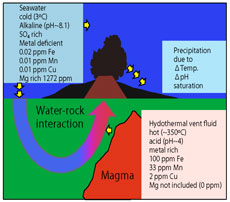
海底熱水循環系は地球システムの重要な駆動要素の一つで,(1)地球内部エネルギーの大放出,(2)大規模鉱床の形成,(3)海水と海洋地殻の化学組成の変化,
(4)熱水生物群集,地下生物圏にとり重要です.昔の拡大系の海底が陸上に衝上したオフィオライトと現在の海底熱水系の試料を研究中です.海洋地殻の成立プロセスの普遍性を明らかにするためには,将来,現在の海底が掘削(「新モホール計画」)が必要です. |
|
 |
|
|
|
<オマーン・キプロスのオフィオライト>
白亜紀に形成された海洋プレートが大陸に乗り上げた岩体で,Sr, O, H,ホウ素(B)の同位体を分析しました.(1)熱水はモホ面まで,(2)オマーンオフィオライトは熱水循環系の流入帯,キプロスオフィオライトは流出帯に位置していました.(3)熱水変質したハンレイ岩はホウ素の大きなシンクです. (4)枕状溶岩のB濃度は,海底風化の期間が長いほど高くなります.(5)海洋地殻のB同位体比は,熱水変質によって約12‰増加しました.
<海底熱水噴出>
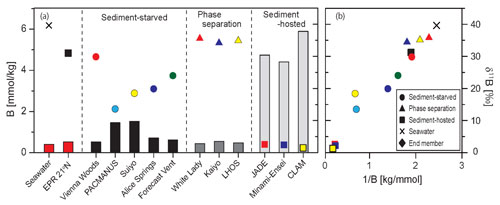
西太平洋の島弧・背弧熱水系で採取された海底噴出熱水について,B,リチウム(Li)の同位体および,B, Li, Rb,Csの濃度分析を行いました.気液分離した熱 水系を除くと,熱水のB,Rb,Cs濃度は中央海嶺に比べて高く,スラブ流体の付加およびマグマの分化によってこれらの元素に富んだ岩石と熱水が反応 したことが示唆されました.
|
|
 |
一方,Li濃度には大きな違いが見られず,岩石にあまり依存しないと考えられました.沖縄トラフでは,熱水がさらに堆積物とも 反応したため,Bおよびアルカリ金属に富むように変化し,その濃度は堆積物に覆われた中央海嶺熱水系よりも 2~5倍高くなりました.
(Yamaoka, Kawahata) |
|
 |
|
|
|
7. 微化石と環境
|
|
|
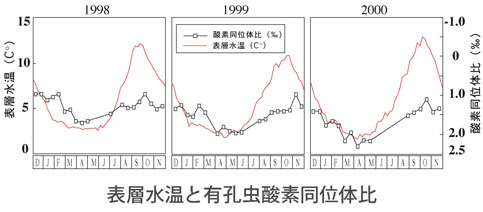
<有孔虫>
有孔虫の研究は黒柳あずみさん(川幡研M,D学生,PDの後,現在東北大助教)と行ってきました.有孔虫は炭酸塩殻化石に生息時の様々な環境を記録しています.私は「有孔虫は微化石の王(女王)様!」と思っています.微化石鑑定をベースとしながらも環境との関連に重点をおいて研究してきました:(1)現代の生態(プランクトンネット,セジメントトラップ),(2)古環境・古気候(化石)の研究,(3)精密飼育実験(温度,酸性度,溶存酸素,硫化水素),(4)遺伝子解析(DNA),(5)殻の化学・同位体比を駆使し,課題に応じて組み合わせて行ってきました.(Kuroyanagi) |
|
 |
|
|
|
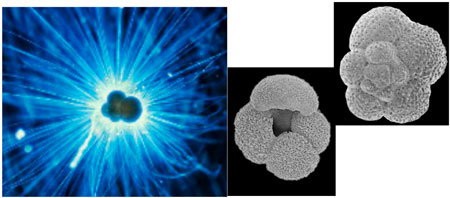
現代の生態:有孔虫研究の重要な基盤データとなる,複数の環境因子を組み合わせた水塊環境や表層・亜表層という僅かな差で異なる環境対応を明らかにしました
精密飼育:溶存酸素濃度をコントロールした浮遊性有孔虫の飼育実験に世界で初めて成功し,これまで考えられていたよりもはるかに高い溶存酸素耐性を持つ事を示しました(海洋酸性化にともなう成果は2ペ-ジを参照)
遺伝子解析(DNA):古環境解析に最もよく用いられるG. ruberという種は,実は遺伝子的には異なる2種が存在し,異なる遺伝子型では生息環境も異なる事を明らかにしました.現在ではこのグループの識別は一般的な手法になってきています.
殻組成:高緯度代表種のN. pachydermaの酸素同位体比と海水温換算式を求め,生息水深及びこれまでは曖昧にされてきた生物効果の複数要因を定量化しました.
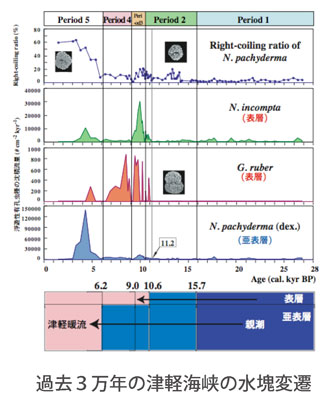
化石(PALEO):前項の「現在の生態」から得られた結果に着目して,過去約3万年の下北半島沖海底堆積物を解析し,最終氷期に孤立していた日本海が,太平洋とリンクしていく過程の津軽海峡の水塊の動きを当時の気候変動とともに明らかにしました.
飼育と化石の融合解析:白亜紀には,何度か海洋無酸素事変(OAE)が起き,そこで有孔虫が高い絶滅・種分化率を示すと言う事が従来報告されてきましたが,前述の飼育実験の結果からそれに疑問を抱き,詳細な解析を行い,無酸素環境が必ずしも直接的な絶滅に至らないないことを示しました. |
|
 |
|
|
|
<放散虫・珪藻>
古生代・中生代・新生代の栄養塩を正確に復元した事例は未だありません.放散虫の殻は古生代には厚く,現在は薄くなってしまいました.この原因は,栄養塩のケイ酸は劇的に下がってきたことに求められます.それを証明した研究はありません.硝酸は脱窒も含めて複雑,リンは適当な同位体がなく,Si(ケイ素)同位体分析法を現在開発中です.チャートを含めて珪質の殻の同位体から顕生代の栄養塩の定量復元をしたいと考えています.(Manaka) |
|
 |
|
|
|
学生と歩んだ研究の成果(修論・博論と国際誌への誌上発表)
山岡香子(2006FY修論,2009FY博論,新領域創成科学研究科長賞)*Yamaoka, K., Kawahata, H., Gupta, L.P., Ito, M. and Masuda, H. (2007) Thermal stabilities of amino acids in siliceous ooze under alkaline hydrothermal conditions. Organic Geochemistry, 38, 1897-1909.
*Ito, M., Yamaoka, K., Masuda, H., Kawahata, H., and Gupta, LP., (2009) Thermal stability of amino acids in biogenic sediments and aqueous solutions at seafloor hydrothermal temperatures. Geochemical Journal, 43, 331- 341.
*Yamaoka,K., Ishikawa, T., Matsubaya, O. Ishiyama, D., Nagaishi, K., Hiroyasu, Y., Chiba, H. and Kawahata, H. (2012) Boron and oxygen isotope systematics for a complete section of oceanic crustal rocks in the Oman ophiolite. Geochim. Cosmochim. Acta, 84,543?559.
三島真理(2007FY修論,2009FY博論,飛級短期終了) *Mishima, M., Kawahata, H., Suzuki, A., Inoue, M., Okai, T. and Omura, A. (2009) Reconstruction of Paleoenvironment at 16 ka in the last glacial period by Faviidae coral collected from Ryukyus. Journal of Quaternary Science, doi:10.1002/jqs.1268.
*Mishima, M., Suzuki, A., Nagao, M., and Kawahata, H. (2010) Abrupt shift toward cooler condition in the Earliest 20th Century Detected in a 165-year Coral Record from Ishigaki Island, Southwestern Japan. Geophysical Research Letters, , 37, L15609, doi:10.1029/2010GL043451.
牛江裕行(2008FY修論,2011FY博論) *Ushie, H., Kawahata, H., Suzuki, A., Murayama, S. and Inoue, M. (2010) Enhanced riverine carbon flux from carbonate catchment to the ocean: a comparative hydrogeochemical study in Ishigaki and Iriomote Islands, southwestern Japan. Journal Geophysical Research, 115, G02017.
*Ushie, H., Matsumoto, K. (2012) The role of shelf nutrients on glacial-interglacial CO2: A negative feedback. Global Biogeochemical Cycle, DOI: 10.1029/2011GB004147.
*Hikami, M., Ushie, H., Irie, T., Fujita, K., Kuroyanagi, A., Sakai, K., Nojiri, Y., Suzuki, A., and Kawahata, H. (2011) Contrasting ocean acidification responses of calcification between two benthic foraminiferal species M. kudakajimensis and C. gaudichaudii., Geophysical Research Letters, 38, Doi:10.1029/2011GL048501.
吉村寿紘(2009FY修論,2012FY博論) *Yoshimura, T., Kawahata, H., Nakashima, R., Suzuki, A., and Tomioka, N. (2010) Oxygen and carbon isotope records of cultured freshwater pearl mussel Hyriopsis sp. shell from Lake Kasumigaura, Japan. Journal of Paleolimnology, 43, 437-448, doi: 10.1007/s10933-009-9341-8.
*Yoshimura, T., Tanimizu, M., Inoue, M., Suzuki, A., Iwasaki, N. and Kawahata, H. (2011) Mg isotope composition of hermatypic and deep-sea corals. Analytical & Bioanalytical Chemistry, 401, 2755-2769.
*Izumida, H., Yoshimura, T., Suzuki, A., Nakashima, R., Ishimura, T., Yasuhara, M., Inamura, A., Shikazono, N. and Kawahata, H. (2011) Biological and water chemistry controls on Sr/Ca, Ba/Ca, Mg/Ca and d18O profiles in freshwater pearl mussel Hyriopsis sp. Paleogeography Paleoclimatology Paleoecology, 309, 298-308.
*Yoshimura, T., Izumida, H., Nakashima, R., Ishimura, T., Shikazono, N., Kawahata, H., Suzuki, A. (2015) Stable carbon isotope values in dissolved inorganic carbon of ambient waters and shell carbonate of the freshwater pearl mussel (Hyriopsis sp.) Journal of Paleolimnology, DOI: 10.1007/s10933-015-9834-6.
*Yoshimura, T., Tamenori, Y., Suzuki, A., Nakashima, R., Iwasaki, N., Hasegawa, H. and Kawahata, H. (2013) Element profile and chemical environment of sulfur in a giant clam shell: insights from μ-XRF and X-ray absorption near-edge structure . Chemical Geology, 352, 170-175, DOI:10.1016/j.chemgeo.2013.05.035.
*Yoshimura, T., Tamenori, Y., Iwasaki, N., Hasegawa, H., Suzuki, A. and Kawahata, H. (2013) Magnesium K-edge XANES spectroscopy of geological standards. Journal of Synchrotron Radiation, 20,734-740, doi:10.1107/S0909049513016099.
小崎沙織(2009FY修論) Iguchi, A., Ozaki, S., Nakamura, T., Inoue, M., Tanaka, Y., Suzuki, A., Kawahata, H. and Sakai, K. (2011) The effect of acidified seawater on coral calcification and symbiotic algae of a massive coral Porites australiensis. Marine Environmental Research, 73, 32-36.
荒岡大輔(2010FY修論,2013FY博論,新領域創成科学研究科長賞) *Araoka, D., Yokoyama, Y., Suzuki, A., Goto, K., Miyagi, K., Miyazawa, K., Matsuzaki, H., and Kawahata, H. (2013) Tsunami recurrence revealed by Porites coral boulders in the southern Ryukyu Islands, Japan. Geology, 41, 919-922, doi:10.1130/G34415.1.
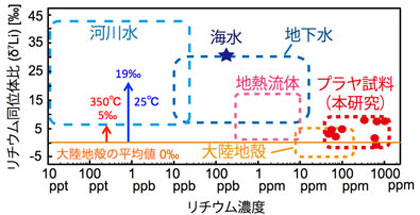
*Araoka, D., Kawahata, H., Takagi, T., Watanabe, Y., Nishimura, K. and Nishio, Y. (2013) Lithium and strontium isotopic systematics in playas in Nevada, USA: constraints on the origin of lithium, Mineralium Deposita, 39, 371-379.
*Araoka, D., Inoue, M., Suzuki, A., Yokoyama, Y., Edwards, R.L., Cheng, H., Matsuzaki, H., Kan, H., Shikazono, N. and Kawahata, H. (2010) Historic geohazard events confirmed by high-resolution U/Th dating of emerged massive Porites boulders at Ishigaki Island in the Ryukyus, Japan. Geochemistry Geophysics Geosystems. , 11, Q06014, doi:10.1029/2009GC002893.
*現在,投稿中(熱水Li同位体)
新免浩太郎(2011FY修論) *Ohki, S., Irie, T., Inoue, M., Shinmen, K., Kawahata, H., Nakamura, T., Kato, A., Nojiri, Y., Suzuki, A., Sakai, K., R. van Woesik (2014) Calcification responses of symbiotic and aposymbiotic corals to near-future levels of ocean acidification. Biogeosciences, 10, 1?8,doi:10.5194/bg-10-1-2013.
松倉誠也(2011FY修論) *Yamaoka, K., Matsukura, S., Ishikawa, T., and Kawahata, H. (2014) Boron isotope geochemistry of the hydrothermally altered oceanic crust and ore deposit from the Troodos ophiolite, Cyprus. Chemical Geology, in press.
氷上 愛(2011FY修論, 2014FY博論,新領域創成科学研究科長賞) *Hikami, M., Ushie, H., Irie, T., Fujita, K., Kuroyanagi, A., Sakai, K., Nojiri, Y., Suzuki, A., and Kawahata, H. (2011) Contrasting ocean acidification responses of calcification between two coral reef benthic foraminiferal species Marginopora kudakajimensis and Calcarina gaudichaudii., Geophysical Research Letters, 38, Doi:10.1029/2011GL048501.
*現在,投稿中(酸素・炭素同位体比)
林 恵里香(2011FY修論) *Hayashi, E., Suzuki, A., Nakamura, T., Iwase, A., Ishimura, T., Iguchi, A., Sakai, K., Okai, T., Inoue, M., Araoka, D., Murayama, S. and Kawahata, H. (2013) Growth-rate influences on coral climate proxies tested by a multiple colony culture experiment. Earth Planetary Science Letters, 362, 198-206.
石川大策(2011FY修論) *Inoue, M., Ishikawa, D., Miyaji, T., Yamazaki, A., Suzuki, A., Yamano, H., Kawahata, H., and Watanabe, T. (2014) Evaluation of Mn and Fe in coral skeletons (Porites spp.) as proxies for sediment loading and reconstruction of 50 yrs of land use on Ishigaki Island, Japan. Coral Reefs, 33, 3630373.
眞中卓也(卒論,2012FY修論) *Manaka, T., Ushie, H., Araoka, D., Yasuhara, M., Inamura, A., Suzuki, A. and Kawahata, H. (2013) Rapid alkalization in Lake Inawashiro, Fukushima, Japan: implications for future changes in the carbonate system of terrestrial waters. Aquatic Geochemistry,19, 281-302, DOI 10.1007/s10498-013-9195-6.
*現在,投稿中(バングラデシュ河川の炭素循環)
森 千晴(2012FY修論) *Tanaka, Y., Iguchi, A., Inoue, M., Mori, C., Sakai, K., Suzuki, A., Kawahata, H. and Nakamura (2013) Microscopic observation of symbiotic and aposymbiotic juvenile corals in nutrient-enriched seawater. Marine Pollution Bulletin, 68, 93-98.
洪 恩松(2012FY卒論) *Yamaoka, K., Hong, E., Ishikawa, T., Gamo, T., and Kawahata, H. (2015) Boron isotopic geochemistry of vent fluids from arc/back-arc seafloor hydrothermal systems in the western Pacific. Chemical Geology, in press.
山下宗佑(卒論) *Kawahata, H., Yamashita, S., Yamaoka, K., Okai, T., Shimoda, G., and Imai, N. (2014) Heavy metal pollution in Ancient Nara, Japan, during the eighth century. Progress in Earth and Planetary Science, 1, 1-15, doi:10.1186/2197-4284-1-15.
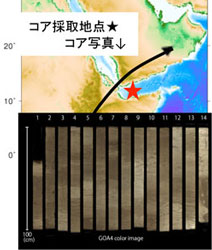
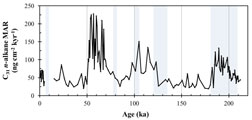
伊左治雄太(卒論,2014FY修論) *Isaji, Y., Kawahata, H.,Ohkouchi, N., Murayama, M., and Tamaki, K. (2015) Terrestrial environmental changes around the Gulf of Aden over the last 210 kyr deduced from the sediment n-alkane record: Implications for the dispersal of Homo sapiens. Geophysical Research Letter, in press.,
*現在,投稿中(ホモ・サピエンスのアフリカ脱出とイエメン沖のアジアモンスーンの変動)
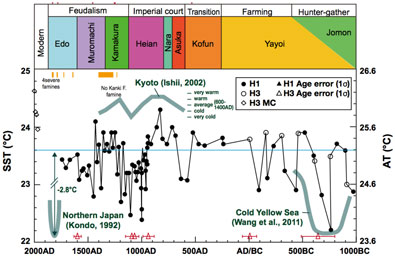
松岡めぐみ(2012FY修論) *現在,投稿中(西日本における過去3000年間の気温復元)
大谷壮矢(卒論,2013FY修論) *現在,投稿中(東南アジア河川の炭素循環)
以下はこちらの都合で遅れていてすみません.
福嶋彩香(卒論,2011FY修論) 近未来投稿準備中(中生代の二酸化炭素分圧推定).
東 賢吾(卒論,2012FY卒論) 近未来投稿準備中(湿原における水環境)
戸上亜美(卒論,2012FY修論) 近未来投稿準備中(大阪における過去3000年間の気温復元)
篠塚 恵(2013FY修論) 近未来投稿準備中(沖縄県地下ダムにおける炭素循環)
小川菜々子(卒論) 投稿準備中(東京・江戸における過去3000年間の気温復元)
修論出版予定なし(島田和明:2008FY,渡辺陽子:2008FY,野崎莉代:中途退学,佐藤愛希子:2009FY,松田直也:2009FY)
上記以外在学中:名越丹理,朝倉 純,Wang, Quan, 平島崇誠,太田雄貴,大澤里美
PDF(授業などで使用した題材も含めて資料を読みやすい形でも掲載しています.)
PDF11. 最新研究テーマ紹介
 11 [クリックしてください.Pdfファイル (15.8MB) が読めます.] 11 [クリックしてください.Pdfファイル (15.8MB) が読めます.]
PDF21. 以前研究テーマ紹介(2014年まで掲載分)
21 [クリックしてください.Pdfファイル (888KB) が読めます.]
PDF3. 考古・歴史時代環境テーマ紹介
PDF31. 平城京都市環境(奈良,東大寺大仏)
31 [クリックしてください.Pdfファイル (6.8MB) が読めます.]
PDF32. 三内丸山遺跡環境
32 [クリックしてください.Pdfファイル (828KB) が読めます.]
PDF33. 縄文時代のまとめ
33 [クリックしてください.Pdfファイル (4MB) が読めます.]
PDF41. 現代の環境と資源問題(今世紀の課題)
41 [クリックしてください.Pdfファイル (1.1MB) が読めます.]
大学院生・若手研究者を歓迎します.この分野の探究に意欲ある若手研究者(博士研究員(PD)),および大学院生(M, D)も歓迎しています.大学院受験の場合には,理学系研究科・地球惑星科学専攻・地球生命圏科学グループ(なお,当研究室を受験する時には,志望欄の「地球生命圏科学グループ」に○をつけてください.(http://www.eps.s.u-tokyo.ac.jp/)
また,ご関心のある方は,どうぞ気楽にメール等(kawahata@aori.u-tokyo.ac.jp)で連絡下さい.また,研究室の見学なども可能です. |
|